BERTON Vincent
GEOFFROY Eric |
Maîtrise de Biologie des Populations et des Ecosystèmes
Université Claude Bernard Lyon 1
Automne 1997
|
Eco-Ethologie
SYSTEMES D’APPARIEMENT
CHEZ LE DAIM (Dama
dama)
Etude du groupe du Parc de la Tête
d’Or

|
Introduction
Introduction
Matériel et
Méthode
Phénologie du
Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
Résultats
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de
l'espace
Discussion
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
Conclusion
Références
|
La sélection naturelle va dans
le sens de la conservation des individus les plus adaptés. Lorsqu'un nouveau
comportement apparaît, si il permet d'améliorer la fitness et (ou) le succès
reproducteur, il se maintiendra, à condition qu'il soit génétiquement régit.
Ainsi, au fil des générations, la compétition (intra ou interspécifique) se
fera sur des critères de plus en plus pointus.
D'après Darwin, la sélection
sexuelle se superpose à la sélection naturelle
et, dans des cas extrêmes, des individus peu adaptés à leur
environnement peuvent fixer leurs gènes si des critères leur permettent
d'avoir une abondante progéniture.
Différentes stratégies peuvent
être élaborées pour l'accès à la reproduction. Plusieurs se rencontrent
chez le daim : elles peuvent être liées au territoire ou pas. En milieu
naturel, les populations peuvent être amenées à se déplacer périodiquement
pour les ressources alimentaires. Dans ces conditions, il n'y a pas de
territorialité. Les populations sont alors organisées en escorte (mâle
suivant un groupe de femelles dans l'intention de copuler),harem (le mâle est
plus ou moins intégré au groupe de femelles) ou groupe hiérarchisé (groupe
de plusieurs femelles et mâles, ces derniers étant dominés par un individu
reproducteur). Ces deux derniers types de stratégie sont fréquents en milieu
à végétation basse (Braza et al.,1986)
Pour des populations plus sédentaires,
les formations se font en postes (permanents ou temporaires, représentent des
territoires non contigus défendus par des mâles solitaires et comportent éventuellement
des ressources alimentaires, le poste temporaire n'étant défendu que quelques
heures dans la journée) ou en leks (les mâles défendent des territoires
contigus, dépourvus de ressource alimentaire). Les formations en leks sont généralement
liées à un fort couvert végétal (Apollonio et
al., 1992).
En dehors de la période de
reproduction, les daims vivent généralement en grandes hardes de femelles avec
leur progéniture, les mâles étant à part, par petits groupes. C'est vers la
deuxième quinzaine d'octobre que débute la période du rut. Elle donne lieu à
un regroupement des deux sexes et
une augmentation de l'agressivité entre les mâles. Les affrontements
aboutissent à une hiérarchisation qui permettrait au mâle dominant d'accéder
aux femelles. Les mâles font alors tout pour se faire repérer des femelles,
s'exposant autant à elles qu'aux prédateurs. Il y a également des risques de
blessure lors des combats et le coût énergétique pour s'assurer une
descendance peut être énorme. Les femelles sont aussi actrices de la sélection
sexuelle. Si elles veulent avoir une progéniture de qualité et plusieurs générations
de descendants, elles doivent choisir des mâles intéressant, d'autant qu'elles
n'ont qu'un petit par an.
A partir d'observations de la
population captive du Parc de la Tête d'Or, parc à très forte densité, on
pourra voir si le système d'appariement est identique à ceux des milieux
naturels. On tentera de décomposer la mise en place de la hiérarchie. On vérifiera
ensuite si la dominance est liée à l'agressivité et si c'est elle qui régit
le succès reproducteur et d'autres facteurs comme le maquillage et
l'occupation de l'espace.
|
| MATERIEL ET METHODE
Introduction
Matériel et
Méthode
Phénologie du
Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
Résultats
Discussion
Conclusion
Références
|
Le groupe de daims du parc de la Tête d’Or est
issu d'une vingtaine de géniteurs, introduits au siècle dernier. Il est
constitué d’environ 150 animaux captifs, certainement génétiquement proche
bien que fitness et fécondité restent élevées (seul la taille des nouveaux nés
parait inférieure à celle de populations sauvages), logés dans un enclos
rectangulaire de 2 hectares. Le sol est herbeux et quelques zones sont
pauvrement plantées. Les promeneurs peuvent, comme nous, longer le parc sur
trois de ses faces. Sur le parc, les animaux disposent d’un abreuvoir, d’un
râtelier à fourrage garni en permanence et d’un abri sommaire pour la pluie
(qu’ils occupent rarement). Des aliments frais (morceaux de légumes et
fruits) ainsi que des granulés leur sont distribués de bon matin.
Phénologie du rut
A partir d’études antérieures
relatant la période à laquelle les comportements reproducteurs ont lieu (vers
la deuxième moitié d’octobre) nous déterminons que c’est début octobre
qu'il faut commencer les observations. Des photos récentes (les bois étant
renouvelés chaque année) permettent de différencier les mâles adultes
(caractérisés entre autres par leur goitre développé et leurs bois à large
empaumure) qui sont «baptisés ». Leur reconnaissance individuelle est
basée sur des signes particuliers, essentiellement la forme et l’asymétrie
des bois qui sont très variables.
La première étape
d’observation est l’entraînement à la reconnaissance des daims et prend au
moins deux heures.
La deuxième, à partir du 9
novembre et par sessions de 2 heures, trois fois par semaine, est le relevé de
la distribution spatiale des mâles connus (les adultes), tous les quarts
d’heure, sur un plan schématique et découpé en huit zones de repérage.
Parallèlement, tous les
comportements agressifs sont placés sur le plan et notés avec la première
lettre du nom des antagonistes. On relève également le type de comportement
agonistique (entraînement, poursuite, combat ou agression de sub-adultes). Les
observations, éparses au début (une fois la première semaine, deux fois la
seconde), s’intensifient en s’approchant du rut. Lorsque arrivent les
montes, elles sont à leur tour notées avec le nom de l'effecteur. Il en sera
également ainsi pour les copulations lorsque celles-ci commenceront et, ceci
indiquant le rut, l’observation en continu sur la phase diurne débutera. Le
temps passé à chaque observation sera noté afin de transformer les données
en fréquences horaires pour chaque type d'activité.
|
|
Matériel et
Méthode
Phénologie du
Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
|
Hiérarchie
Les interactions agonistiques de
chaque individu sont reportées dans des matrices de dominance (une par semaine
et une comportant la totalité des observations) faisant apparaître les combats
perdus en ligne et ceux gagnés en colonne (Annexes XXX, matrices 95, 96, 97).
Ces matrices permettent d'établir
les diagrammes de flux qui mettent en évidence les relations de dominance entre
les mâles et le nombre d'interactions qu'ils ont eu.
Afin de disposer de valeurs
quantitatives concernant la hiérarchie, l'indice de Clutton-Brock est calculé:
iCB(x)=B+Sb+1 / (L+Sl+1)
B=nombre
d'individus contre lesquels l'individu x a gagné plus de fois qu'il n'a perdu
Sb=nombre
d'individus inférieurs aux B individus, jamais inférieurs à x, un même
individu ne pouvant être deux fois au numérateur.
L=
nombre d'individus contre lesquels l'individu x a perdu plus de fois qu'il n'a
gagné
Sl=nombre
d'individus supérieurs aux B individus, jamais supérieurs à x, un même
individu ne pouvant être deux fois au dénominateur.
Cet indice tient compte du
nombre de daims hiérarchiquement inférieurs (daims vaincus ainsi que de ceux
qu'ils dominaient) et du nombre de daims hiérarchiquement supérieurs (ceux
directement supérieurs ainsi que de ceux qui les dominent).
Les indices sont calculés sur
la matrice totale de cette année et des deux précédentes. Les indices peuvent
être confrontés au diagramme de flux, au succès reproducteur, au
maquillage,...
|
|
Matériel et
Méthode
Phénologie du
Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
|
Evolution du maquillage
Le maquillage de la face et du
plastron (pelage enduit d'une croûte boueuse) est relevé avec la date et le
nom de chaque individu.
|
|
Matériel et
Méthode
Phénologie du
Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
|
Occupation de l'espace
Les données concernant la
localisation des mâles sont regroupées:
-en quatre zones pour voir l'évolution
de leur fréquentation
-en deux zones pour tester la
différence de distribution de la totalité de la harde puis de dominants et de
dominés avant et pendant le rut.
|
| Résultats
Introduction
Matériel et
Méthode
Résultats
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de
l'espace
Discussion
Conclusion
Références
|
Phénologie
du rut
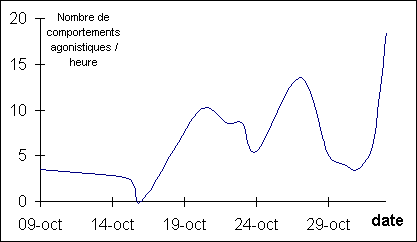
Figure 1:
Evolution de la fréquence des comportements agonistiques
On note une
augmentation de la fréquence des comportements agonistiques jusqu'au 27 Octobre
puis une diminution en fin de pré-rut. Ces données contiennent également les
agressions de sub-adultes qui représentent environ 70% des comportements
agonistiques le 27 Octobre. Pendant la phase du rut, une augmentation de la
fréquence des combats et des poursuites a été observée mais n'a pas été
quantifiée (aucun relevé le 1 Novembre et un seul de deux heures le 2
Novembre).
Le graphe
suivant permet de faire la distinction entre les trois types de comportements
agonistiques concernant les adultes:
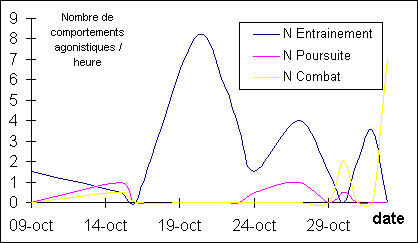
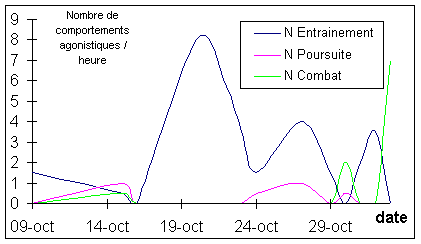
Figure
2: Evolution de la fréquence de chaque type
de comportements agonistiques
La fréquence des entraînements
est la plus élevée le 20 Octobre, celle des poursuites le 27 et celle des
combats le 30 et pendant la phase du rut (non quantifiée).
Figure
3: Evolution de la fréquence des montes et des copulations
Le nombre de
montes augmente brusquement entre les 24 et 27 Octobre et reste élevé lors du
rut. Les copulations commencent le 2 Novembre, onze sont comptabilisées sur la
journée et sont le fait de sept mâles différents.
|
|
Résultats
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de
l'espace
|
Hiérarchie
Les matrices de
dominance (cf. Annexe 1) rendent bien compte de l'augmentation de l'agressivité
puisque sur l'ensemble du pré-rut (du 6 au 26 Octobre), 55 comportements
agonistiques ont été observés contre 91 pour le rut (du 27 Octobre au 2
Novembre).
Les diagrammes
de flux (Figures 4; 5; 6 p.5,7 et 8) montrent que Kaki est dominant la dernière
semaine alors qu'il a très peu interagi les semaines précédantes. En effet,
lors du pré-rut, il n'est qu'en huitième position et, les mâles qui semblent
s'imposer sont Gaston et Ding-dong. Pendant cette période, bien que Kaki ait
très peu interagi, il n'a jamais été dominé. En revanche, lors du rut, il
s'impose devant Ding-dong et Gaston. Sur la totalité de la période, l'ordre
hiérarchique est le suivant:
K>D>G>F>E>J>A>M>H>B>I>L>Q>C>P
On note une
discordance entre cette hiérarchie, basée uniquement sur les interactions
dominantes des mâles et celle établie d'après l'indice de Clutton-Brock
(1991) (calculs détaillés et classements en Annexe 2) sur
l'ensemble de la période d'étude, qui est présentée ci-dessous. Cette
discordance apparaît à partir du quatrième individu:
K>D>G>J>L>B>F>E>A>M>I>H>C>Q>P
Les
extrémités des hiérarchies sont très similaires avec les deux types de
classement. C'est pour les individus intermédiaires que l'ordre varie. Ainsi,
Fluck "chute" de la quatrième à la septième place alors que Léo
passe de la douzième à la cinquième place.
Afin d'évaluer
la liaison entre l'indice de dominance des mâles et leur succès reproducteur,
le graphe suivant a été élaboré, mettant les deux variables en relation,
pour les études de 1995, 1996 et 1997:
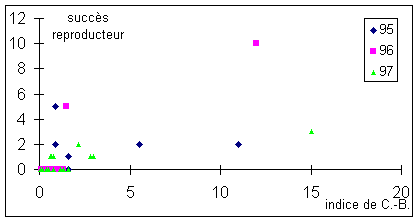
Figure 7: Succès reproducteur en fonction de l'indice de dominance
Pour
les trois années, les individus ayant les plus faibles indices de Clutton-Brock
n'ont généralement pas accès à la reproduction. En revanche, il y a
globalement une augmentation du nombre de copulations avec celle de l'indice.
Cette année,
l'ordre du succès reproducteur est: K>J>D=E=G=I=M
(Annexe 3), ce qui ne correspond pas vraiment à la hiérarchie observée.
J&B a un succès reproducteur supérieur à ce que sa place dans la
hiérarchie laisserait présager.
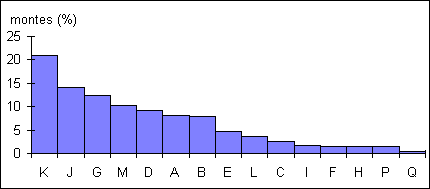
Figure
8: Pourcentage de montes assurées par chaque mâle les 1 et 2 Novembre
L'ordre
des individus classés en fonction de la quantité de montes n'est pas fortement
lié à l'ordre hiérarchique. Si Kaki assure à lui seul un cinquième des
montes, Fuck, bien placé dans la hiérarchie se retrouve parmi les derniers.
|
|
Résultats
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de
l'espace
|
Evolution
du maquillage
Le tableau de
résultats concernant l'évolution du maquillage (cf. annexe 4) peut être
transformé en histogrammes. Avant tout, précisons que les relevés du 16
Octobre ne seront pas pris en compte car il pleuvait et que la présence d'un
maquillage de la poitrine a été notée pour chaque daim alors qu'il ne
s'agissait que de saleté. De même, les relevés du 31 Octobre et du 1°
Novembre ne semblent pas très logiques et ne seront donc pas pris en compte car
du jour au lendemain, les treize daims maquillés à la poitrine ne le sont
plus.
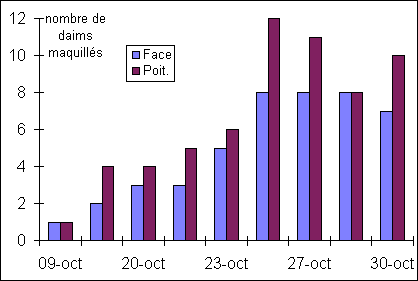
Figure
9: Evolution du nombre de daims maquillés
On
constate qu'il y a globalement une augmentation du nombre de daims maquillés
jusqu'au 27 Octobre, où il y a stabilisation. Le maquillage de la face et de la
poitrine augmentent en même temps mais, les individus maquillés à la poitrine
ne sont jamais moins nombreux que ceux ayant la face maquillée.
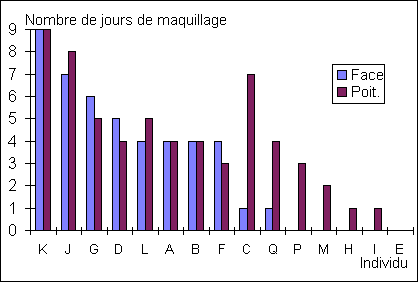
Figure
10: Nombre de jour de maquillage avant le rut pour chaque daim
Cette figure montre que Kaki est
resté maquillé le plus longtemps. Il a de plus été le premier maquillé (Annexe 4)
|
|
Résultats
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de
l'espace
|
Occupation
de l'espace
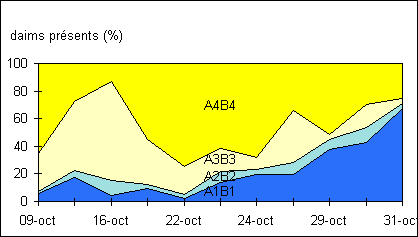
Figure
11: Evolution de la répartition des mâles sur quatre portions du parc
L'observation
indépendante de l'occupation de chaque portion n'apporte pas beaucoup
d'éléments sinon que A1B1 est de plus en plus fréquenté, au détriment de
A3B3. L'occupation de A2B2 reste constante et faible. En observant les portions
deux à deux (A4B4,A3B3 et A2B2,A1B1),la moitié contenant le poste se dépeuple
progressivement au profit de l'autre (comparaison pré-rut /rut :c²obs=155.6>>c²theo5%=3.84).
Tableau I:
Comparaison de la distribution spatiale de trois dominants et trois dominés
| |
A1B1 |
A2B2 |
A3B3 |
A4B4 |
|
|
| |
obs
|
théo
|
obs
|
théo
|
S
|
c²partiels
|
| Kaki
|
12
|
21,38
|
78
|
68,6
|
90
|
5,39
|
| Ding-dong
|
19
|
20,66
|
68
|
66,3
|
87
|
0,18
|
| Gaston
|
15
|
20,19
|
70
|
64,8
|
85
|
1,75
|
| Quetch
|
16
|
19,48 |
66
|
62,5
|
82
|
0,81
|
| Pince
|
15
|
18,05
|
61
|
57,9
|
76
|
0,68
|
| Cornu
|
42
|
19,24
|
39
|
61,8
|
81
|
35,3
|
| S
|
119
|
|
382
|
|
501
|
44,1 =
c²
|
L'hypothèse
nulle Ho d'égalité des distributions est refusée avec un risque de 5%. Les
plus fortes contributions au c² sont
dues à Cornu qui est beaucoup plus en A1B1A2B2 qu'attendu et à Kaki, plus en
A3B3A4B4 qu'attendu si la distribution était aléatoire. Les autres individus
ont des distributions similaires.
|
| Discussion
Introduction
Matériel et
Méthode
Résultats
Discussion
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
Conclusion
Références
|
Le mâle possédant un
territoire de reproduction le délimite par des signaux olfactifs (phéromones),
sonores (réements) et visuels. L'entrée d'un autre mâle sur le territoire
entraîne automatiquement un affrontement qui aboutira à l'expulsion d'un mâle.
Le groupe étudié au Parc de la Tête d'Or semble présenter un système
d'appariement intermédiaire entre le poste et le groupe hiérarchisé : nous
verrons qu'une hiérarchie s'établit entre les mâles du groupe pour la
possession du poste.
Phénologie du rut
En dehors de la période de
reproduction, il y a relativement peu de comportements agonistiques ; ceux-ci
augmentent globalement jusqu'au rut (figure 1) afin de déterminer quel mâle
aura accès aux femelles. Trois types de comportements agonistiques se succèdent
(figure 2). Cette succession se traduit par un accroissement de la violence des
affrontements. Les premiers à se manifester (à partir du 16 octobre) sont les
entraînements et ce sont aussi les plus nombreux. Ils permettent une mise en
place grossière de la hiérarchie qui évitera des combats plus violents entre
daims de force très différente. Les poursuites (à partir du 24 octobre),
beaucoup moins nombreuses, représentent le deuxième type de comportement
agonistique et exigent une dépense énergétique supérieure. Elles sont
suivies des combats (le 30 octobre) qui s'intensifient avant la détermination
du poste (le 1 novembre) et se maintiennent ensuite. Ces combats, parfois très
violents, déterminent toujours un vainqueur et établissent le haut de la hiérarchie
de la harde.

Kaki
a l’assaut d’un concurrent trop ambitieux
(2
novembre)
Les montes (figure 3) débutent
approximativement en même temps que les poursuites et précèdent le rut de
quelques jours. Elles seraient responsables du déclenchement de l'œstrus des
femelles et constitueraient une étape clef de la mise en place du rut. Celui-ci
débute avec les premiers accouplements (2 novembre) qui coïncident avec
l'ovulation des biches. Les observations n'ayant pu se poursuivre après le 2
novembre, les données concernant le succès reproducteur sont incomplètes
cette année et la durée du rut reste inconnue. Les années précédentes, les
accouplements se sont étalés sur trois (1995) à sept jours (1996) (Annexes 5
et 6).
Le rut s'achève après les
copulations, lorsque l'émission phéromonale des femelles désormais fécondées
s'atténue. Les mâles cessent alors de les courtiser et de s'affronter.
|
|
Discussion
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
|
Hiérarchie
Sur la harde du parc, plusieurs
hiérarchies peuvent être établies selon qu'on se base sur le nombre
d'interactions dominantes ou sur l'indice de Clutton-Brock. Les diagrammes de
flux (Figures 4 ; 5 et 6) présentent les animaux classés selon le nombre
d'affrontements gagnés. L'ordre ne correspond pas forcément à la hiérarchie
car, des animaux ayant gagné de nombreux combats mais en ayant perdu davantage
se retrouvent bien classés alors que leur taux de victoires peut être faible .
Le diamètre des cercles, proportionnel à la fréquence des interactions, est,
pour ces individus, plus important que pour les individus hiérarchiquement supérieurs.
De plus, ce classement n'identifie pas les antagonistes. Si un mâle domine
plusieurs fois un autre individu, il sera classé au même niveau qu'un mâle
ayant dominé peu de fois de nombreux individus.
L'indice de Clutton-Brock fait
la distinction entre les deux cas précédents en tenant également compte des
relations eues par les animaux dominants l’individu considéré et par ceux
qu’il domine. Quel que soit le mode de classement adopté, les extrémités
sont identiques, ce qui montre que, pour ces individus, les dominances sont
clairement établies. Ainsi, Kaki est incontestablement le mâle dominant. En
considérant les diagrammes de pré-rut et rut (figure 4 et 5), on observe que
la dominance de Kaki ne s'est affirmée qu’au moment du rut, c’est à dire
lorsque ses intérêts reproducteurs étaient réellement en jeu. Pendant le pré-rut,
il n'occupait que la huitième place (avec seulement trois interactions
dominantes) et s'est imposé lors de la phase ultime du rut (42 affrontements
gagnés contre seulement 2 perdus). Plusieurs hypothèses concernant l'établissement
soudain de sa dominance sont émises. Les animaux, étant en relation en
permanence, connaissent peut-être la supériorité de Kaki et ne le défient
pas (un suivi individuel des comportements agonistiques depuis la naissance
permettrait de vérifier cette hypothèse). Il se peut également que les
affrontements impliquant Kaki se déroulent à notre insu, en dehors des périodes
d'observations (notamment la nuit). Enfin, on peut soupçonner une stratégie
consistant à s'effacer alors que les autres s'affrontent, permettant ainsi une
économie d'énergie. Cette dernière hypothèse est confortée par le fait que
Kaki a dû affronter Pince et Momo
(hiérarchiquement bas), sans être inquiété par les plus forts, occupés à
se "battre" entre eux. Ce sont peut-être les phéromones émises par
les femelles en œstrus qui accroissent son agressivité quand arrive le rut.
Corrélativement à sa position
hiérarchique, Kaki a le plus fort succès reproducteur (3 copulations, Annexe
3). Les copulations du mâle dominant ont normalement lieu sur le poste, zone
dont il interdit l'accès aux autres mâles et sur laquelle il rée et répand
des phéromones. Les femelles s'y présentent et adoptent des postures caractéristiques
: toutes dans le même sens, debout ou couchées, tête dressée, dos arrondi.
C'est la femelle qui décide du moment de l'accouplement en barrant le chemin au
mâle qui s'active sur son poste. Il lui renifle les parties génitales afin de
connaître son état ovulatoire. S’il est positif, il y a saillie, sinon, il
la stimule par une monte sans copulation. Ce comportement permet au mâle
dominant, qui a accès à un grand nombre de femelles, de réserver sa semence
à celles qui sont fécondables.

Kaki
rée sur le poste alors qu’une femelle rumine, en position caractéristique
Etant donné le nombre
d’animaux sur le parc (environ 150 bêtes, dont 15 mâles adultes), on aurait
pu s’attendre à la formation d’un deuxième poste. L’espace restreint et,
par conséquent, la densité
importante empêchent cette formation. Lors du rut, les animaux passifs ou dominés
se replient en A1B1 et n’auraient plus qu’un refuge limité si un deuxième
poste s’établissait. La distance réduite d’une extrémité à l’autre du
parc et l’absence d’obstacle visuel mettraient les mâles aux postes en
relation visuelle, auditive et olfactive en permanence, ce qui les pousseraient
à combattre pour éliminer la concurrence.
Les mâles sub-dominants peuvent
eux aussi avoir accès aux femelles mais, généralement, dans une moindre
mesure et en dehors du poste. Momo a cependant copulé une fois sur le poste le
2 novembre, lors d'une trêve de la part de Kaki. Cinq autres mâles (G, D, J, E
et I) ont également assuré quelques copulations ce même jour en divers lieux
(Annexe 3). Il arrive que des sub-adultes parviennent à s'accoupler en trompant
la vigilance des femelles occupées à boire ou à manger. Ces comportements
sournois s'apparentent à des viols et, on peut penser qu'ils aboutissent à la
fécondation d'un certain nombre de femelles, ce qui permettrait d'expliquer
qu'ils aient été sélectionnés et qu'ils se soient génétiquement maintenus
au fil des générations.
Le succès reproducteur semble
globalement lié à l'indice de Clutton-Brock (figure 7). Les individus ayant un
succès reproducteur nul ont tous un indice relativement faible (inférieur à
deux). En revanche, la réciproque n'est pas vraie puisque des individus tels
que Momo ont pu copuler. Il semblerait donc que la dominance ne soit pas le seul
critère de choix du partenaire sexuel par les femelles. Le mâle dominant peut
refuser de prendre une femelle s’il ne la sent pas fécondable ou qu'elle ne
l'intéresse pas. Cette femelle peut se rabatte sur un autre mâle pour assurer
sa fécondation. Le choix de ce mâle peut se faire d'après :
- son classement hiérarchique
(D ; G et J bien que J ait assuré une copulation de plus que les deux autres),
idée renforcée par le fait que tous les individus ayant un indice supérieur
à deux ont eu accès aux copulations sur les trois dernières années.
- son obstination contre le mâle
dominant (J, qui l'affronte neuf fois).
- l'originalité de ses bois
(raquettes planes et peu découpées de D ; bois "veloutés" de M ou
asymétriques de E et G). On peut citer, pour ce dernier cas, le concept de
l'avantage du mâle rare.
Les mâles dominés ne refusent
pas cette occasion de disperser leurs gènes tout comme les femelles qui ont
plus intérêt à avoir une descendance génétiquement moyenne que pas de
descendance du tout.
La quantité de montes n'est pas
complètement liée à l'ordre hiérarchique. En revanche, elle reflète assez
bien le succès copulatoire. Soit l'ordre des montes (figure 8) est déterminé
par la cote des mâles, soit cet ordre est du au nombre de tentatives des mâles
envers les femelles qui cèdent parfois.

Une
copulation
|
|
Discussion
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
|
Evolution du maquillage
Le maquillage est visuel et
odorant. Pour le constituer, le mâle gratte le sol et urine sur la terre mise
à nu et sur son plastron à l'aide de son pinceau pénien. Il se couche ensuite
et s'enduit la poitrine de boue mélangée à l'urine chargée en phéromones prépuciales.
Cette boue peut également servir au maquillage de la face, sur laquelle les phéromones
préorbitales sont également étalées.
Le nombre de jours de maquillage
(figure 10) est corrélé linéairement (R²=0.84, en ne prenant que la face,
sur les onze premiers individus) au nombre de montes, donc l'ordre des mâles
classés suivant leur durée de maquillage correspond à celui du succès
reproducteur. Ainsi, plus un mâle est maquillé tôt, plus il copulera. Il est
peu probable que ces variables sont directement liées, car cela supposerait que
les femelles mémorisent les dates de maquillage. En revanche, il est possible
que la durée de maquillage avant le rut et le succès reproducteur soient
fortement liés à la hiérarchie réelle, légèrement différente de celle établie
par l'indice de Clutton-Brock (R² maquillage-indice=0.5).
Si la durée de maquillage
(figue 10) détermine la hiérarchie, cela suppose que les mâles connaissent
leur place avant même la période de reproduction. On peut en fait supposer que
la date de premier maquillage facial correspond aux "prétentions" des
individus, qui étaient justifiées cette année (il serait bon d'avoir des données
concernant les années précédentes pour savoir si c'est un accident). S'il était
possible de reconnaître les mâles d'une année sur l'autre, nous pourrions
savoir si la prétention des mâles est liée à la hiérarchie de l'année précédente.
Comme pour l'épinoche, qui
considère, en période de reproduction, les poissons de couleur rouge comme des
concurrents mâles de son espèce, le maquillage facial chez le daim est un
signe que l'individu du même sexe a des prétentions hiérarchiques. Pourtant,
cet ornement ne déclenche pas d'agressivité car Kaki, jusqu'au 27 octobre, n'a
interagi que trois fois (pendant nos observations) malgré sa cohabitation avec
sept mâles maquillés sur le parc (figure 9). Le maquillage peut aussi servir
à attirer l'attention des femelles
|
|
Discussion
Phénologie
du Rut
Hiérarchie
Evolution du maquillage
Occupation de l'espace
|
Occupation de l'espace
En raison de la taille réduite
du parc par rapport à la mobilité des animaux, ceux-ci peuvent passer d'une
extrémité à l'autre très rapidement et l'absence d'obstacle visuel leur
permet de surveiller l'ensemble de l'enclos à partir de n'importe quel point.
De ce fait, les animaux accourent tous dès que les gardiens ou, plus fréquemment,
les promeneurs les nourrissent. Ceci perturbe notablement la distribution qui
est en moyenne plus importante en A4B4 (figure 11), où le nourrissage est le
plus commode et où la mangeoire et l'abreuvoir sont disposés. Il s'avère que
c'est également près des limites de ce secteur que le poste sera établi, en
A3B3.
En s'approchant du rut, A3B3 est
progressivement délaissé (figure 11), ainsi que A4B4 dans une moindre mesure
(l’intérêt alimentaire reste élevé), au profit de A1B1, partie très calme
du parc : peu d'affrontements, peu de visiteurs et beaucoup de femelles avec
leurs petits. La fréquentation de A2B2 est constante et faible. C'est une zone
sans arbre, ne semblant pas présenter d’intérêt pour les animaux. Il y a
une baisse significative (c²=156.5>>3.84)
de la fréquentation de la moitié A3B3A4B4 entre la phase de pré-rut et celle
du rut –(l’utilisation du test de c²
n’est pas rigoureuse car les échantillons ne sont pas indépendants)-. La
comparaison de distributions de trois individus dominants et trois individus
dominés (Tableau I) permet de voir qu’il n’y a pas de différence notable
entre les individus inférieurs au mâle dominant (sauf pour Cornu qui occupe
indifféremment les deux moitiés du parc). Le seul a être nettement plus représenté
en A3B3A4B4 qu’attendu est Kaki qui chasserait les autres de cette moitié. Il
n'est pas seul responsable de cette "désertion" puisque des travaux
d'aménagement de l’enclos sont venus perturber les observations à compter de
la semaine du 20 octobre, ce qui introduit un biais supplémentaire. En fait,
les années précédentes, il y avait au contraire une augmentation de la fréquentation
autour du poste jusqu’à la phase ultime du rut, à partir de laquelle le
secteur était plus délaissé (à cause de la territorialité du dominant). On
peut alors supposer que
l’influence des travaux sur le comportement des animaux n’est pas négligeable.
De plus, l’empiétement du chantier sur l’aire A3B3 réduit sa surface et
par conséquent sa fréquentation. Ceci peut être la cause de la forte baisse
de fréquentation de A3B3 entre le 16 et le 22 octobre (figure 11).
Le poste était établi au même emplacement
en 1995 et 1996 mais, cette année, pour la mise en place du chantier,
l’abreuvoir a été déposé sur cette zone. Le retard dans la détermination
du poste (le 30 octobre cette année, entre le 23 et le 27 en 1996 et 1995) est
attribué a ces perturbations ainsi qu’au stress causé par les travaux. En
plus d’être en retard, le poste était très peu marqué et de limites imprécises,
aux alentours de l’abreuvoir. Les causes de la localisation stable du poste au
fil des années ne sont pas connues. On peut supposer une mémorisation visuelle
de l’emplacement des années précédentes, à moins que le poste soit déterminé
par un cadre général (Apollonio et al.), 1992)(zone dégagée, distance des ressources alimentaires,
…) ou encore une rémanence d’odeurs phéromonales. L’incertitude peut être
levée en déplaçant la couche superficielle du sol en dehors de la période de
reproduction ou en déplaçant les animaux dans des parcs aux cadres différents.
Le 2 novembre, (seul jour de phase ultime de rut
observé), Kaki était très irrégulier dans la défense du poste. Parfois,
d’autres mâles pouvaient pénétrer sur le poste sans être inquiétés alors
qu’à d’autres moments, le passage à une dizaine de mètres entraînait des
poursuites. On peut penser qu’il s’agit soit d’un comportement normal :
le mâle, qui s’est peu alimenté et beaucoup dépensé les jours précédents,
aurait des baisses de vigilances passagères, soit, que cela est lié aux
perturbations exceptionnelles de cette année et Kaki n’a pas la motivation
suffisante pour assurer une défense soutenue du poste qui est peu fréquenté
par les femelles.
L’absence des femelles au
poste a contribué au retard des premiers accouplements (2 novembre contre 27
octobre en 1996 et 31 octobre en 1995) si on ne prend pas en compte la
copulation du 27 octobre qui pourrait être une erreur d’appréciation vu sa
précocité. En fait, cette copulation a lieu à la même date que les premières
saillies des années précédentes et
le mâle qui l’a fait est peut-être le seul qui ne soit pas déréglé par
les perturbations inhabituelles.
|
| Conclusion
Introduction
Matériel et
Méthode
Résultats
Discussion
Conclusion
Références
|
L’analyse des comportements
est délicate car les observations ont été faites à des horaires variables.
Les animaux ayant un rythme nycthéméral (phases de manducation, de repos ou de
digestion, de forte activité, …), il serait préférable de les étudier à
horaire constant pour voir l’évolution des comportements dans le temps sans
que les variations quotidiennes interviennent. Les comportements agonistiques,
par exemple, varient certainement moins entre deux journées consécutives
qu’entre l’aube (ou le crépuscule) et le milieu de journée.

|
| Références
Introduction
Matériel et
Méthode
Résultats
Discussion
Conclusion
Références
|
-
Brazza et al., Rutting Behaviour of Fallow Deer,
1986
-
Apollonio et al., To lek or not to lek :
mating strategies of male fallow deer, 1992
-
Clutton-Brock, Materetension, harasment, and the
evolution of ungulates leks, 1991
-
Le monde
animal en treize volumes, vol. XIII, editions Stauffacher, 1972
|
|